|
Banneling
Geregistreerd: 18 februari 2003
Berichten: 26.968
|
Citaat:
Oorspronkelijk geplaatst door CUFI
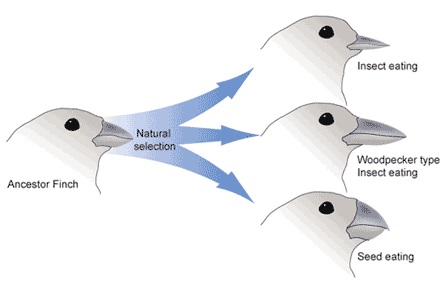
Hoe namen ze waar dat vissen muteerden tot een andere soort?
Leg eens concreet in eigen woorden uit met bronvermelding.
|
Via duizenden overgangsvormen natuurlijk. 
En de waarneming bestaat natuurlijk uit het vinden en opgraven van de fossielen van die overgangsvormen in de juiste sedimentaire lagen.
Citaat:
Samenvatting van het gekende gewervelde fossiel bestand
(We beginnen met primitieve kaakloze vissen.)
Overgang van primitieve kaakloze vissen naar haaien, roggen en vleten
Laat Siluur – eerste kleine haaiachtige tandachtige uitstekels.
Vroeg Devoon – eerste herkenbare haaientanden, duidelijk afgeleid van schubben.
LEEMTE: Merk op dat de eerste, zeer, zeer oude sporen van haaiachtige dieren zo fragmentarisch zijn dat we er niet veel gedetailleerde informatie uit kunnen halen. Bijgevolg weten we niet welke kaakloze vis de feitelijke voorouder was van de vroege haaien.
Cladoselachium (laat Devoon) – Prachtige vroege haaifossielen, gevonden in graafwerken tijdens de bouw van snelwegen tussen staten in de V.S. Waarschijnlijk geen directe voorouders van haaien, maar het geeft een merkwaardig beeld van de vroege anatomie van de haai, tot de spierweefsels toe!
Tristychius & gelijkaardige hybodonten (early Mississippium) -- Primitieve proto-haaien met vinnen met een brede wijze die voor de rest sterk lijken op die van de haai.
Ctenacanthus & gelijkaardige ctenacanthiden (laat Devoon) -- Primitieve, trage haaien met haaiachtige vinnen met een brede basis & vinvormige uitsteeksels. Waarschijnlijk voorvaderlijk voor alle moderne haaien, roggen en vleten. Fragmentarische vinvormige uitsteeksels (Triassic) – van meer geavanceerde haaien.
Paleospinax (vroeg Jurassic) – Meer geavanceerde kenmerken zoals een losse bovenkaak, maar behoudt primitieve ctenacanthide kenmerken zoals twee dorsale ruggengraten, primitieve tanden, etc.
Spathobatis (laat Jurassic) -- Eerste proto - rog.
Protospinax (laat Jurassic) – Een zeer vroege haai/vleet. Nadien de eerste heterodonten, hexanchiden, & baby haaien verschijnen (laat Jurassic). Andere haaiengroepen dateren uit het Krijttijdperk of Eoceen. De eerste echte haaien zijn gekend vanaf het Boven Krijttijdperk.
Een aparte afstamming loopt van de A separate lineage ctenacanthiden via Echinochimaera (laat Mississippium) en Similihari (laatPennsylvanium) naar de moderne ratvis.
Overgang van primitieve kaakloze vissen naar beendervissen.
Boven Siluur – eerste kleine schubben gevonden.
Leemte: Hier opnieuw zijn de eerste sporen zo fragmentarisch dat we de feitelijke voorvader niet kunnen identificeren.
Acanthodians(?) (Siluur) – Een intrigerende groep van stekelige vissen met gelijkenissen met vroege beendervissen.
Palaeoniscoiden (bijvoorbeeld. Cheirolepis, Mimia; vroeg Devoon) -- Primitieve beendervissen met straalvinnen die aan de basis liggen van de grote meerderheid van de levende vissen. Zware acanthodianachtige schubben, acanthodianachtige schedel, en een grote rudimentaire ruggengraat.
Canobius, Aeduella (Carboon) -- Latere paleoniscoiden met kleinere, meer geavanceerde kaken.
Parasemionotus (vroeg Triassic) -- "Holosteaanse" vis met gewijzigde kaken maar nog steeds heel wat primitieve kenmerken. Bijna exact halverwege de late paleoniscoiden & eerste teleosten. Merk op: de meeste van deze vissen leefden in seizoensgevoelige rivieren en hadden longen. Ik herhaal: de longen ontwikkelden zich het eerst in vissen.
Oreochima & gelijkaardige pholidophoriden (laat Triassic) – De meest primitieve teleosten, met lichtere schubben (bijna rondschubbig), gedeeltelijk verbeende ruggenwervels, meer ontwikkelde wangen en kaken.
Leptolepis & gelijkaardige leptolepiden (Jurassic) – Meer geavanceerd met volledig verbeende wervels & cirkelvormige schubben. De leptolepiden uit het Jurassic verspreidden zich tot de moderne teleosten (de omvangrijke, succesvolle groep van vissen die vandaag bijna volledig dominant zijn). Longen transformeerden in longblaasjes.
Alen & sardines dateren van het late Jurassic, zalm van het Paleoceen & Eoceen, karper van het Krijttijdperk, en de grote groep van stekelvormige teleosten van het Eoceen. De eerste leden van vele van deze families zijn gekend en behoren tot de familie van de leptolepiden (noteer het inherente classificatie probleem!).
Overgang van primitieve beendervissen naar amfibieën
Weinig mensen realiseren zich dat de overgang van vis naar amfibie er geen was van water naar land. De overgang van vinnen naar poten vond immers plaats in het water. De allereerste amfibieën lijken benen en poten te hebben ontwikkeld om rond te snellen op de bodem in het water, zoals sommige moderne vissen doen, en niet om op het land rond te wandelen (zie Edwards, 1989). Dit aquatisch potenstadium betekende dat vissen niet snel hoefden te veranderen, de gewichtdragende lidmaatmusculatuur hoefde niet goed ontwikkeld te zijn en de axiale musculatuur hoefde helemaal niet te veranderen. Recent gevonden gefragmenteerde fossiele poten uit het midden Boven Devoon (zie lager), ondersteunen dit idee van "aquatische poten" stadium. Uiteindelijk, natuurlijk, trokken de amfibieën inderdaad het land op. Dit hield in dat het bekken steviger aan de ruggengraat moest bevestigd worden, en dat de schouder van de schedel diende gescheiden te worden. De longen waren geen probleem, aangezien longen een kenmerk van antieke vissen zijn en reeds aanwezig waren.
Paleoniscoiden opnieuw (e.g. Cheirolepis) – Deze antieke beenderbissen stonden aan de wieg van zowel de modern straalvinvissen (hoger vermeld), en ook de vissen met lobvormige vinnen.
Osteolepis (midden Devoon) – Een van de vroegste crossopterygiaanse vissen met lobvormige vinnen, die nog steeds enkel kenmerken delen met de longvissen (de andere vissen met lobvormige vinnen). Ze hadden gepaarde vinnen met een potenschikking en grote ledematenbeenderen, die in staat waren te buigen aan de “elleboog”, en ze hadden een vroege amfibieachtige schedel en tanden.
Eusthenopteron, Sterropterygion (midden laat Devoon) -- Vroege rhipidistiaanse vissen met lobvormige vinnen ruwweg halverwege de vroege crossopterygiaanse vissen en de vroegste amfibieën. Eusthenopteron is het best gekend van een ongewoon volledig fossiel dat voor het eerst gevonden werd in 1881. De schedel is erg amfibieachtig. Sterke amfibieachtige ruggengraat. Vinnen die sterk lijken op de poten van vroege amfibieën wat betreft het algemene ontwerp van de belangrijkste beenderen, de aanhechting van de spieren, en de beenderprocédés, met een tetrapodachtige tetrahedrale humerus, en tetrapodachtige elleboog- en kniegewrichten. Maar er zijn geen waarneembare “tenen”, enkel een stel identieke vinstralen. De verhoudingen van lichaam en schedel lijken eerder op die van een vis.
Panderichthys, Elpistostege (midden laat Devoon, ongeveer 370 Mjg) -- Deze"panderichthyiden" zijn erg tetrapodachtige vissen met lobvormige vinnen. Verschillend van de Eusthenopteron, lijken deze vissen ook echt op tetrapods in hun algemene verhoudingen (afgeplatte lichamen, dorsaal geplaatste oogkassen, frontale beenderen! in de schedel, rechte staarten, etc.) ze hebben merkwaardig pootachtige vinnen.
Gefragmenteerde ledematen en tanden uit het midden van het Laat Devoon (ongeveer 370 Mjg), mogelijk behorend tot de Obruchevichthys – Ontdekt in 1991 in Schotland, zijn dit de vroegst gekende tetrapod overblijfselen. Het opperarmbeen is voor het grootste gedeelte tetrapodachtig maar behoud enkele viselementen. De ontdekker Ahlberg (1991), zei: "Het [het opperarmbeen] is meer tetrapodachtig dan om het even welk opperarmbeen van een vis, maar het mist de karakteristieke vroeg-tetrapod 'L-vorm'...dit blijkt een primitief, visachtig kenmerk te zijn....hoewel het scheenbeen duidelijk tot een poot behoort, verschilt het opperarmbeen voldoende van het vroege tetrapod patroon om het onzeker te maken of het aanhangsel tenen droeg dan wel een vin. Op het eerste gezicht lijkt de combinatie van zulke twee extremen binnen hetzelfde dier hoogst onwaarschijnlijk op functionele gronden. Als echter tetrapod ledematen zich ontwikkelden voor beweging in het water eerder dan op het land, zoals recent werd gesuggereerd, kan zo’n morfologie perfect werkbaar zijn.
LEEMTE: Idealiter willen we natuurlijk een volledig skelet uit het midden van het laat Devoon, en niet enkel fragmenten van ledematen. Niemand heeft er tot nog toe een gevonden.
Hynerpeton, Acanthostega, en Ichthyostega (laat Devonian) – Een beetje later was de vin-naar-poot overgang complete, en we hebben een stel viervoetige fossielen die duidelijk poten hadden. De meest volledige zijn Ichthyostega, Acanthostega gunnari, en de nieuw beschreven Hynerpeton bassetti (Daeschler et al., 1994). (Er zijn ook andere geslachten gekend via gefragmenteerde fossielen.) Hynerpeton is de vroegste van deze drie geslachten (365 Mjg), maar is op sommige punten meer geavanceerd; de twee andere geslachten behielden langere tijd meer visachtige kenmerken dan dit het geval was met de Hynerpeton afstamming.
Labyrinthodonten (eg Pholidogaster, Pteroplax) (laat Devoon/vroeg Mississippium.) – Deze grotere amfibieën hebben nog steeds enkele viskenmerken van de icthyostegiden, zoals schedelbeenpatronen, ingewikkeld tandbeen, de aanwezigheid & het patroon van grote palatale slagtanden, het gewricht van een visschedel, stukken van een kieuwstructuur tussen de wangen en de schouder, en de vertebrale structuur. Maar ze missen heel wat andere viskenmerken : de fijne stralen in de start zijn verdwenen, de wervels zijn sterker en grijpen in elkaar in, de nasale doorgang voor het inademen is goed gedefinieerd.
Meer info over deze eerst gekende amfibieën uit het Laat Devoon: Acanthostega gunnari was sterk visachtig, en recent ontdekten Coates & Clack (1991) dat hij nog steeds inwendige kieuwen had! Ze zeiden: "Acanthostega lijkt visachtige inwendige kieuwen te hebben bewaard en een open operculaire kamer voor het gebruik bij aquatische ademhaling, wat impliceert dat de eerste viervoetige amfibieën geen uitsluitende landdieren waren....Het behoud van visachtige inwendige kieuwen bij een viervoeter uit het devoon doet het traditionele onderscheid vervagen tussen viervoeters en vissen... Dit biedt verdere ondersteuning aan de suggestie dat unieke viervoetige kenmerken zoals ledematen met tenen zich eerst ontwikkelden voor het gebruik in het water eerder dan voor rond te wandelen op het land. " Acanthostega had ook een opmerkelijk visachtige schouder en voorste lidmaat. . Ichthyostega was ook bijzonder visachtig door het behoud van een visachtige gevinde staart, een permanent lateraal lijnsysteem en een rudimentaire ruggengraat. Geen van beide dieren hebben lang kunnen overleven op het land.
Coates & Clack (1990) ontdekten ook recentelijk de eerste goedbewaarde poten van de Acanthostega (voorpoot gevonden) en Ichthyostega (achterpoot gevonden). (Poten van de Hynerpeton zijn niet bekend.) De poten waren veel meer vinachtig dan men verwacht had. Men had altijd verondersteld dat ze vijf tenen hadden aan elke poot, zoals bij alle moderne viervoeters. Dit was een raadsel aangezien de vinnen van vissen met lobvinnen niet schijnen voort te bouwen op basis van een vijftenig plan. Het blijkt nu dat de voorpoot van de Acanthostega acht tenen had, en dat de achterpoot van de Ichthyostega zeven tenen had, beide poten het uitzicht gevend van een korte, krachtige zwempoot met talrijke “teenstralen” gelijkaardig aan vinstralen. Het enige wat je met een lobvin moet doen om het te maken tot zo een veeltenige poot, is hem krullen, de vinstralen rond het uiteinde van het lidmaat windend. In feite is dit precies de manier waarop de poten zich ontwikkelen vanuit een gebogen lidmaatknop bij de larven van amfibieën. (Lees ook Goulds essay bmet betrekking tot dit onderwerp, "Eight Little Piggies".) De ontdekkers zeiden (Coates & Clack, 1990): "De morfologie van de ledematen van Acanthostega en Ichthyostega suggereren een aquatische levenswijze, compatibel met een recente vaststelling van de vis-viervoeter. The dorsoventraal gecomprimeerde lagere bootbeenderen van de Ichthyostega lijken sterk op die van een cetaceaanse [walvis] pectorale zwemvin. Een bijzondere, van beenderen slecht voorziene massa ligt voorafgaand en aangrenzend aan de tenen, en lijkt een versterking te zijn voor het voorste uiteinde van dit peddelachtige lidmaat." Coates & Clack ontdekten ook dat de voorpoot van de Acanthostega niet kon voorwaarts buigen aan de elleboog, en dat hij dus niet in een gewichtdragende positie kon gebracht worden. Met andere woorden, deze “poot” fungeerde nog steeds als een horizontale vin. De achterpoot van Ichthyostega kan best op dezelfde manier gefungeerd hebben, hoewel zijn voorpoot gewicht kon dragen. Functioneel gezien zijn deze beide dieren geen volkomen amfibieën; zij leefden in een vis/amfibie tussenniche met poten die nog steeds gedeeltelijk fungeerden als vinnen. Hoewel zijn waarschijnlijk geen voorouders zijn van de latere viervoeters, tonen Acanthostega & Ichthyostega beslist aan dat de overgang van vis naar amfibie mogelijk is!
Hynerpeton, daarentegen, had waarschijnlijk geen inwendige kieuwen en beschikte reeds over een goed ontwikkelde schoudergordel, hij kon zijn voorpoot krachtig opheffen en terugtrekken en hij beschikte over sterke spieren die zijn schouders verbonden met de rest van zijn lichaam. (Daeschler et al., 1994). De ontdekkers van de Hynerpeton denken dat, aangezien hij als eerste de sterkste voorpoten had, hij wel eens de feitelijke voorvader kan zijn geweest van alle hierna komende vierpotige landdieren, terwijl Acanthostega and Ichthyostega een zijtak kunnen zijn geweest die gelukkig waren met hun hoofdzakelijk aquatische niche en daar bleven.
Samengevat, de allereerste amfibieën (momenteel slechts gekend van fragmenten) waren waarschijnlijk bijna volledig aquatisch, hadden zowel longen als inwendige kieuwen gedurende heel hun leven, en snelden rond onder water met zwemvinachtige, veeltenige poten die niet veel gewicht droegen. Verschillende afstammelingen van amfibieën begonnen ofwel hun achterpoten ofwel hun voorpoten naar voren te buigen zodat de poten gewicht konden dragen. Eén lijn van afstammelingen (Hynerpeton) droeg gewicht op alle vier de poten, ontwikkelde sterke lidmaatgordels en spieren en begon zich snel op het land te bewegen.
Overgangen tussen amfibieën
Temnospondylen, bijvoorbeeld Pholidogaster (Mississippium, ongeveer 330 Mjg) – Een groep van grote labrinthodonte amfibieën, een overgangsfase tussen de vroege amfibieën (de ichthyostegiden, hierboven beschreven) en latere amfibieën zoals such rhachitomes en anthracosaurus. Ze stonden waarschijnlijk ook aan de wieg van de modern amfibieën (de Lissamphibia) via deze keten van zes temnospondyle geslachten, een progressieve wijziging vertonend van het verhemelte, gebit, oor, en de pectorale gordel met een gestage vermindering van de lichaamsgrootte (Milner, in Benton 1988). Merk nochtans op dat de tijdstippen door elkaar lopen , hoewel ze alle dateren uit het Pennsylvanium en vroeg Perm. Ofwel ontstonden sommige van de "Permi" geslachten vroeger, in het Pennsylvanium (hoogst waarschijnlijk), en/of sommige van deze geslachten zijn “neven”, geen directe voorouders (ook zeer waarschijnlijk) .
Dendrerpeton acadianum (vroeg Penn.) -- 4-tenige hand, rechte ribben
Archegosaurus decheni (vroeg Perm) – Verdwenen intertemporale
Eryops megacephalus (laat Penn.) -- Occipitale condylus gesplitst in 2
Trematops spp. (laat Perm) – Middenoor zoals bij moderne amfibieën
Amphibamus lyelli (midden-Penn.) -- Dubbele occipitale condylussen, zeer smalle ribben
Doleserpeton annectens of misschien Schoenfelderpeton (allebei vroeg Perm) -- Eerste pedicellate tanden! (een klassiek kenmerk van moderne amfibieën
Van daar springen we naar het Mesozoicum
Triadobatrachus (vroeg Triassic) -- een proto -kikker, met een langere romp en een veel minder gespecialiseerd heupbeen, de staart is nog steeds aanwezig (maar zeer kort) .
Vieraella (vroeg Jurassic) – eerst bekende echte kikker.
Karaurus (vroegJurassic) – eerst bekende echte salamander.
Naamloze proto-anthracosaurus – beschreven door Bolt et al., 1988. Dit dier combineert primitieve kenmerken van palaeostegalianen (bijvoorbeeld temnospondylachtige gewervelde dieren) met nieuwe anthracosaurusachtige kenmerken. Anthracosaurussen waren een groep van grote amfibieën waarvan men denkt dat ze uiteindelijk geleid hebben tot de reptielen. Gevonden in een nieuwe Lager Carboon site in Iowa, van ongeveer 320 Mjg.
Lagomorfen (haasachtigen)
Barunlestes de mogelijke Aziatische knaagdier/ lagomorf voorouder
Mimotoma (paleoceen). Een konijnachtig dier, gelijkaardig aan Barunlestes, maar met een tandenformule als van een konijn, veranderingen in de gezichtsbeenderen, en slechts één laag email op de snijtanden (niet als de knaagdieren). Zoals konijnen had het twee bovenste snijtanden, maar de tweede snijtand is nog groot en functioneel, terwijl deze bij het moderne konijn klein is. Chuankuei-Li et al. (1987; zie ook Szalay e.a.., 1993) denken dat dit de eigenlijke voorouder is van Mimolagus, zie volgende.
Mimolagus (laat eoceen.) Beschikt over meer lagomorfachtige eigenschappen, zoals een speciale emaillaag, mogelijk dubbele bovenste snijtanden, en grote voorkiezen.
Lushilagus (midden-laat-Eoceen. Eerste echte lagomorf. Tanden zeer gelijkend op Mimotoma, en de moderne konijnentanden en hazentanden kunnen gemakkelijk uit deze tanden zijn afgeleid.
Hierna verschenen de eerste moderne konijnen in het oligoceen.
Gekende soort-soort transities in lagomorfen:
De midden tertiaire lagomorf Prolagus vertoont een zeer mooie “chronocline” (graduele verandering over de tijd) verandering van de ene soort naar de andere. Gingerich (1977) zegt: “ In Prolagus toont een zeer compleet fossiel record een opmerkelijke maar continue en geleidelijke reorganisatie van de morfologie van de kroon van de voorkies in een enkele evolutielijn”.
Lundelius e.a.. (1987) vernoemen transities in pleistocene konijnen, vooral bij Nekrolagus tot Sylvilagus, en van Pratilepus tot Aluralagus. Merk op dat deze beide transities genuslijnen overschrijden. Zie ook de paper over de lagomorf in Chaline (1983). Sommige van deze transities werden beschouwd als “plotse verschijningen” tot de tussenliggende fossielen werden bestudeerd, waarbij talrijke transitionele individuen aan het licht kwamen.
Condylarthen, de eerste hoefdieren
Protungulatum (laatste krijt). Transitioneel tusen de eerste placentazoogdieren en de condylarthen (primitieve, kleinhoevige dieren). Deze vroege, eenvoudige insectivoorachtige kleine zoogdieren hadden één nieuwe ontwikkeling: hun kaaktanden hadden malende oppervlakken in plaats van eenvoudige, gepunte toppen.
Zij waren de eerste herbivore zoogdieren. Al hun andere kenmerken zijn algemeen en primitief – eenvoudige plantigrade klauwpoten met vijf tenen, alle tanden aanwezig (3:1:4:3) zonder tussenopeningen, alle beenderen van de ledematen aanwezig en niet verbonden, puntige schedel, smalle en kleine hersenen, oogkas niet gesloten. In de tijdspanne van enkele miljoenen jaren splitsten de condylarthen zich in licht verschillende soorten met iets verschillende tanden, zoals oxyclaeniden (de meest primitieve), triisodontinen, en phenacodonten (beschreven in andere secties). De eerste verschillen vergrootten doorheen de tijd naarmate de soorten verder en verder uit elkaar gingen. Uiteindelijk resulteerde dit in zulke verschillende dieren als walvissen, miereneters, en paarden. Het is interessant te zien hoe de vroege soorten condylarthen op elkaar geleken, in contrast tot hoe verschillend hun afstammelingen uiteindelijk langzamerhand werden. Paleontologen geloven dat dit een klassiek voorbeeld is van hoe “hogere taxa” zoals families en ordes ontstaan.
Zegt Carroll (1988, p 105): “ In het geval van de ceteanen (walvissen) en de perissodactylen (paarden etc), werd hun afkomst uit de condylarthen duidelijk gedocumenteerd…. Indien, zoals het aannemelijk lijkt, het mettertijd mogelijk zou zijn om de voorouders van de meeste placentadieren terug te volgen naar het vroege paleoceen, of zelfs het laatste krijt, zullen de verschillen tussen de voorouderlijke vormen klein zijn – potentieel niet groter dan deze die soorten onderscheiden of zelfs populaties binnen soorten. De oorsprong van ordes zal synoniem worden met de oorsprong van soorten of geografische subgroepen. In feite is dit patroon wat men zou verwachten van ons begrip van de evolutie teruggaand tot Darwin. De selectieve krachten verbonden aan de oorsprong van belangrijke groepen zou worden gezien als niet verschillend van deze die leiden tot aanpassing aan zeer licht verschillende omgevingen en levenswijzen. Op basis van een beter begrip van de anatomie en de relatie tussen de vroegste ungulaten (hoefdieren), kunnen we zien dat de oorsprong van de ceteanen en de perissodactylen niet voortkwam uit grote verschillen in hun anatomie en levenswijze maar uit lichte verschillen in hun dieet en manier van bewegen. We merken dit aan het patroon van de bovenkant van de tanden en aan details van de beenderen van carpus (handwortel) en tarsus (voetwortel) (p. 505).
Soort-soort transitie binnen de condylarthen
Het meest voorkomende fossiele zoogdier uit het lager eoceen is een kleine primitieve wezelachtige condylarth, Hyopsodus genaamd. Het was voorheen bekend dat veel zeer verschillende soorten Hyopsodus gevonden waren op verschillende sites, met (bijvoorbeeld) zeer verschillende tandgroottes. In 1976 analyseerde Gingerich de tandgrootte van alle gekende fossielen van Hyopsodus die betrouwbaar en onafhankelijk konden worden gedateerd. Hij stelde vast dat “het patroon van verandering in tandgrootte dat verschijnt er een van een voortdurende graduele verandering tussen soorten, met graduele divergentie volgend op de afscheiding van nieuwe zustergeslachten”. Wanneer de tandgrootte wordt getoond op een tijdsschaal, ziet men dat de enkele soort zich geleidelijk splitst in vier soorten nakomelingen (dit was één van de eerste gedetailleerde en uitgebreide studies over soortvorming).
Tegen 1985 had Gingerich veel meer specimen van Hyopsodus en van verschillende andere eocene condylarthensoorten, zoals Haplomylus. Bijvoorbeeld: “Haplomylus speirianus… werd geleidelijk groter doorheen de tijd, uiteindelijk gestalte gevend aan een nieuwe soort Haplomylus scottianus…. Hyopsodus latidens werd ook groter en vervolgens kleiner, uiteindelijk uitmondend in een nog kleinere soort, Hyopsodus simplex”. Deze analyses waren gebaseerd op honderden nieuwe specimens (505 voor Haplomylus en 869 voor Hyopsodus) afkomstig uit Clarck’s Fork Basin in Wyoming. Opgemerkt dient te worden dat verschillende andere soorten uit dezelfde tijd stilstand toonden (vooral Ectocion, waarvan voordien was gezegd dat hij verandering toonde, maar die in feite bijna hetzelfde bleef) en dat niet alle soortentransities zijn gedocumenteerd. Dus transities worden niet altijd gevonden. Maar soms worden ze wel gevonden.
Cetaceanen (walvissen, dolfijnen)
Tot voor enkele jaren was er nog steeds een grote lacune in het fossiele record van cetaceanen. Er werd gedacht dat zijn voortkwamen uit landlevende mesonychiden die geleidelijk hun achterpoten kwijt waren geraakt en in het water gingen leven. De evolutietheorie voorspelde dat zij door een fase moeten zijn gegaan waarin zij gedeeltelijk in het water leefden maar nog steeds achterpoten hadden, maar er waren geen gekende intermediaire fossielen. Een vloed van recente ontdekkingen uit India en Pakistan (de oevers van de oude Tethys-zee) heeft deze lacune bijna volledig opgevuld. Er zijn nog steeds geen gekende soort-soort transities en de “keten van soorten” is niet compleet. Maar we hebben een gedeeltelijke evolutielijn en zoals voorspeld hebben de nieuwe walvisfossielen poten. (voor besprekingen zie Berta, 1994; Gingerich e.a. 1990, Thewissen e.a. 1994; Discover magazine, Jan 1995; Gould 1994)
Eoconodon of gelijkaardige triisodontine arctocyoniden (vroeg paleoceen). Niet gespecialiseerde condylarthen vrij gelijkend op de vroege oxyclaenide condylarthen, maar met sterke hoektanden (die de eerste vleesetende neiging aantonen), stompe malende kaaktanden en afgeplatte klauwen in plaats van nagels.
Microclaenodon (midden paleoceen). Een transitioneel genus intermediair tussen Epconodon en de mesonychiden, met kiezen die geplaatst staan op velerlei manieren om op voorkiezen te lijken. Meer aangepast aan vleeseten.
Dissacus (midden paleoceen). Een mesonychide (niet gespecialiseerd vleesetend dier uit het paleoceen) met kiezen meer gelijkend op voorkiezen en verschillende andere veranderingen in tanden. Had nog altijd vijf tenen in de voet en een primitieve plantigrade houding.
Hapalodectes of een zeer gelijkaardige mesonychide (vroeg eoceen, rond 55 Ma). - Een kleine mesonychide met zeer smalle scharende kiezen, een uitgesproken gevormde zygomatische boog en eigenaardige beaderde delen tussen de kiezen. Waarschijnlijk een lopend dier dat kon zwemmen door met de poten te peddelen. Hapalodectes zelf is misschien te laat om de voorouder van de walvis te zijn, maar was waarschijnlijk een dicht familielid van de voorouder van de walvis. Zegt Carroll (1988) “ De schedels van eocene walvissen dragen onmiskenbare gelijkenissen met die van primitieve landzoogdieren uit het vroege cenozoïcum. Vroege (walvis)genera hadden een primitieve tandentelling met duidelijk onderscheiden snijtanden, hoektanden, voorkiezen en kiezen met meervoudige wortels. Hoewel de snuit langwerpig is, vertoont de vorm van de schedel gelijkenissen met die van de mesonychiden, vooral Hapalodectes…”.
Pakicetus (vroeg midden eoceen, 52 Ma). Het oudste gekende walvisfossiel. Zelfde schedelkenmerken als Hapalodectes, nog altijd met een zeer aards landoor (tympanisch membraan, geen bescherming tegen drukveranderingen, geen goede onderwatergeluidlokalisatie), en daarom duidelijk geen diepe duiker. Kiezen hebben nog altijd zeer mesonychideachtige toppen, maar andere tanden zijn als die van latere walvissen. Neusgaten nog altijd vooraan de kop (geen spuigat). Walvisachtige schedel en langwerpige kaken. Ledematen niet bekend. Slechts 2,5 m lang. De schedel werd gevonden samen met landfossielen en kan amfibisch geweest zijn, zoals van een nijlpaard.
Ambulocetus natans (vroeg midden eoceen, 50 Ma). Een recent ontdekte vroege walvis, met voldoende ledematen en wervels aanwezig om te zien hoe de vroege walvissen zich voortbewogen op het land en in het water. Deze walvis had vier poten! De voorpoten waren stomp. Achterpoten waren kort maar goed ontwikkeld, met enorme brede voeten die uitstaken als staartvinnen. Had geen echte staartvinnen, enkel een eenvoudige lange staart. Grootte zoals een zeeleeuw. Had nog altijd een lange snuit zonder spuigat. Stapte waarschijnlijk op het land als een zeeleeuw en zwom zoals een zeehond/ otter door te sturen met de voorpoten en zich aan te drijven met de achterpoten. Dus, juist zoals voorspeld, waren deze vroege walvissen erg gelijkend op onze moderne zeeleeuwen. Zij konden zwemmen, maar zij konden nog altijd op het land lopen (Thewissen e.a.., 1994).
Rodhocetus (midden eoceen, 46 Ma). Nog een zeer recente (1993) ontdekking van een fossiele walvis. Had achterpoten die een derde kleiner waren dan die van A. natans. Kon waarschijnlijk nog een beetje “waggelen” op land, maar had tegen deze tijd een sterke staart (aangetoond door massief grote staartwervels) en kon waarschijnlijk in de zee blijven gedurende lange perioden. Neusgaten waren een stukje naar achteren opgeschoven van de top van de snuit weg.
Basilosaurus isis, Protocetes, Indocetus ramani en gelijkaardige walvissen met korte poten uit het midden van het laat eoceen (45-42 Ma). - Na Rodhocetus kwamen verschillende walvissen die nog altijd achterpoten hadden, maar er niet meer op konden lopen. Bijvoorbeeld B. isis (42 Ma) had achterpoten met 3 tenen en een klein restant van de 2e teen (de grote teen ontbreekt geheel). De poten waren klein en moeten onbruikbaar geweest zijn voor voortbeweging maar waren gespecialiseerd voor het voorwaarts zwaaien in een vaste schrijlingse positie. Waarschijnlijk een hulp bij copulatie voor het lange lichaam van deze langlijvige slangachtige walvis. B.isis kan een “neef” geweest zijn van onze moderne walvissen, geen directe voorouder. Een andere recente ontdekking is Protocetes, een iets meer geavanceerde walvis uit het laat eoceen. Deze was ongeveer 3 m lang (zoals een dolfijn), en had nog een primitieve tandenrangschikking, neusgaten aan het einde van de snuit, en een groot bekken aangehecht aan de wervelkolom; ledematen onbekend. Tot slot Indocetus, enkel gekend door fragmentarische overblijfselen, maar deze omvatten een scheenbeen. Deze laat-Eoceen walvissen met poten hadden nog altijd mesonychideachtige tanden, en in feite waren enkele van deze fossielen eerst verkeerd geïdentificeerd als mesonychiden toen enkel de tanden gevonden waren (zie Gingerich e. a. 1990 voor meer info over B. isis).
Prozeuglodon (laat eoceen, 40 Ma). Een andere recent ontdekte walvis, gevonden in 1989. Had bijna de achterpoten verloren maar niet helemaal; droeg nog altijd een paar rudimentaire achterpoten van 15 cm aan zijn 3 meter lange lichaam.
Eocetus en gelijkaardige “archeocete walvissen” uit het laat Eoceen. Deze meer geavanceerde walvissen hebben hun achterpoten volledig verloren, maar houden een schedel en tanden als een “primitieve walvis”, met niet gefuseerde neusgaten. Zij groeiden uit tot een grotere lengte (tot 25 m tegen het einde van het eoceen) en hadden en lang, gestroomlijnd lichaam, zwemvliezen en een kraakbeenachtige staartvin. Het oor was aangepast om onder water te horen. Merk op dat deze fase van aanpassing aan het leven in het water bereikt werd ongeveer 15 miljoen jaar na de eerste mesonychiden die op het land leefden.
Dorudon intermedius – een walvis uit het laat eoceen die waarschijnlijk de voorouder is van de moderne walvis.
In het Oligoceen splitsten walvissen in twee evolutielijnen:
Tandwalvissen:
Agorophius (laat oligoceen). Schedel gedeeltelijk ineengeschoven, maar kaaktanden nog altijd geworteld. Intermediair op vele manieren tussen archaeocetes en latere tandwalvissen
Prosqualodon (laat oligoceen). Schedel geheel verlengd met neusgaten bovenaan (spuigat). Kaaktanden vermeerderd in hoeveelheid maar hebben nog steeds oude toppen. Waarschijnlijk voorouder van de meeste latere tandwalvissen (met mogelijke uitzondering van de Sperm-walvissen).
° Kentriodon (midden mioceen). Schedel verlengd, nog symmetrisch. Lopen in het laat mioceen uit in de moderne dolfijnen en kleine tandwalvissen met asymmetrische schedels.
Balein (tandloze) walvissen:
Aetiocetus (laat oligoceen). De meest primitieve gekende mysticete walvis (= baleinwalvis) en waarschijnlijk de stamgroep van alle latere baleinwalvissen. Had een mysticeteachtig los kaakgewricht en luchtsinus, maar had nog alle tanden.
Mesocetus (midden mioceen) verloor zijn tanden
Moderne baleinwalvissen verschenen voor het eerst in het laat mioceen.
Perissodactyla,(paarden, tapirs, neushoorns)
Hier komen we tot de meest bekende algemene evolutielijn van al, de paardensequentie. Het was de eerste dergelijke evolutielijn die ontdekt werd, in de late jaren 1800, en werd daarom de meest beroemde. Er is een eigenaardig gerucht dat circuleert in de kringen van creationisten dat de paardensequentie op een of andere manier verdacht of verouderd is. Dit is niet zo; het is een zeer goede sequentie die enkel meer gedetailleerd is uitgegroeid en compleet gemaakt over de jaren, voor het grootste gedeelte veranderend door de toevoeging van grote zijtakken. Zoals deze verschillende paleontologen recent hebben gezegd: “ het uitgebreide fossiele record levert een uitstekend voorbeeld van grootschalige evolutionaire verandering op lange termijn.” (Colbert, 1988). “Het fossiele record (van paarden) is een helder verhaal van afkomst met verandering gedurende bijna 50 miljoen jaar, en we weten veel over de voorouders van de moderne paarden “ (Evander, in Prothero en Schoch, 1989, p 125). Al de morfologische veranderingen in de geschiedenis van de Equidae kunnen gestaafd worden door de neo-Darwiniaanse theorie van micro-evolutie: genetische variatie, natuurlijke selectie, genetische afwijking en soortvorming “ (Futuyama, 1986, p 409”). Fossiele paarden leveren inderdaad sluitend bewijs van de evolutionaire theorie (MacFadden, 1988).
Hier dus de opsomming van de paardensequentie.
Loxoplophus (vroeg paleoceen). Een primitieve condylarth met vrij laagkronige kiezen, waarschijnlijk voorouder van de phenacodontide condylarthen.
Tetraclaenodon (midden paleoceen). Een meer gevorderde paleocene condylarth uit de phenacodontide familie, en bijna zeker voorouder van alle perissodactylen (een andere orde). Lange maar niet gespecialiseerde ledematen, 5 tenen aan elke voet (#1 en #5 kleiner). Iets meer efficiënte pols.
LACUNE: Er zijn bijna geen gekende fossielen van persissodactylen uit het laat paleoceen. Dit is eigenlijk een kleine lacune; ze is enkel merkbaar omdat het perissodactyle fossiele record anders zeer volledig is. Recente ontdekkingen hebben duidelijk aangetoond dat de eerste perissodactylen opkwamen in Azië (een weinig bestudeerd continent), dus hopelijk zullen de aan de gang zijnde nieuwe fossielenjachten in Azië deze kleine maar frustrerende lacune vullen. De eerste aanwijzing is al binnen:
Radinskya yupingae (laat paleoceen, China). Een recent ontdekte perissodactylachtige condylarth (McKenna e.a.., in Prothero en Schoch, 1989)
Hyracotherium (vroeg eoceen, ongeveer 55 Ma; voorheen “Eohippus”). Het beroemde “Paard van de dageraad”, een kleine hondachtige perissodactyl, met een gebogen rug, korte nek, omnivore tanden en korte snuit. 4 tenen vooraan en 3 achteraan. Vergeleken met Tetraclaenodon heeft hij langere tenen, samenhangende enkelbeenderen, en lichtjes verschillende tandentoppen. Waarschijnlijk geëvolueerd uit Tetra. Over ongeveer 4-5 miljoen jaar, mogelijk via een Aziatische soort zoals Radinskya. Merk op dat Hyrac. verschilde van andere vroege perissodactylen (zoals de tapir/ neushoorn voorouders), enkel door kleine veranderingen in tandentoppen en lichaamsgrootte.
Hyracotherium vassacciense (vroeg eoceen). De specifieke soort waaruit in het bijzonder waarschijnlijk de Equiden voortkwamen.
Orohippus (midden eoceen – 45 Ma). Klein, 4/3-tenig, ontwikkelt tandrichels van een loofeter.
Epihippus (laat eoceen – 45 Ma). Klein, 4/3 tenig, goede tandrichels, loofeter
Epihippus (Duchesnehippus). Een later subgenus met Mesohippus-achtige tanden
Mesohippus celer (laatste eoceen, 40 Ma). Drietenig aan alle poten, loofeter, ietsje groter
Mesohippus westoni (vroeg oligoceen). Een iets latere, meer geavanceerde soort.
Miohippus Assiniboiensis (midden oligoceen). Deze soort splitste zich af van de vroege Mesohippus via cladogenetische evolutie, waarna Miohippus en Mesohippus overlapten gedurende de volgende 4 miljoen jaar. Duidelijk groter, iets langere schedel, gezichtsgroeven dieper en meer uitgebreid, licht verschillende enkelgewricht, variabele extra richel op de tanden van de bovenkaak. In het vroeg mioceen (24 Ma) begon Miohippus snel te specialiseren. Grassen hadden zich pas ontwikkeld en tanden begonnen overeenkomstig te veranderen. Poten etc. begonnen te veranderen om snel te lopen.
Kalobatippus (laat oligoceen). Drietenige loofeter met voet tussen Mio en Para.
Parahippus (vroeg mioceen, 23 Ma). Drietenige loofeter/ grazer, ontwikkelt veervoet. Permanente aanwezigheid van de extra richel die zo variabel was bij Miohippus. Sterkere tandrichels en lichtjes lagere tandkronen
‘Parahippus’ leonensis (midden eoceen ~ 20 Ma). Drietenige loofeter/ grazer met de nadruk op grazer. Ontwikkelt veervoet en hoogkronige tanden
‘Merychippus’ gunteri (midden mioceen, ~ 18 Ma). Drietenige grazer, volledig met veervoeten en met hoogkronige tanden.
Merychippus primus (midden-Mioceen, ~17 Ma). Iets meer gevorderd.
Merychippus ssp uit midden laat mioceen (16-15 Ma). Drietenige grazers, met veervoeten, formaat van een kleine pony. Diversifieerde in alle beschikbare niches van grazers, waaruit tenminste 19 succesvolle drietenige grazers voortkwamen. Zijtenen in verschillende groottes, zeer klein in sommige lijnen. De paardachtige hoef ontwikkelt, beenderen van de poten fuseren. Volledig hoogkronige tanden met dik cement en dezelfde ribbels als Parahippus. De lijn waaruit uiteindelijk Equus voortkwam ontwikkelde als volgt: M. primus, M. sejunctus, M. isonesus (deze laatste twee hadden nog altijd een mix van primitieve, hipparion en equine kenmerken), M. intermontanus, M. stylodontus, M. carrizoensis. Deze laatste twee leken paardachtig, met vrij kleine zijtenen, en hieruit kwam een set grotere drietenige en ééntenige paarden bekend als de “echte equines”. Zonneklaar nietwaar?
KLEINE LACUNE: Het is niet bekend uit welke Merychippussoort (stylodontus? Carrizoensis?) de eerste Dinohippusssoorten voortkwamen (Evander, in Prothero en Schoch, 1988).
Dinohippus (laat mioceen, 12 Ma). Eentenige grazer, met veervoeten. Erg equine voeten, tanden en schedel, met rechtere tanden en kleinere gezichtsgroeven. De eerste was D. spectans, gevolgd door D. interpolatus en D. leidyanus. Een iets latere soort was D. mexicanus, met nog rechtere tanden en nog kleinere gezichtsgroeven.
Equus (Plesippus), ook de “E. simplicidensgroep” genoemd (Plioceen, ~ 4 miljoen jaar). Drie dichtverwante soorten van eentenige veervoetige hoogkronige grazers. Geen gezichtsgroeven en zeer rechte tanden. De grootte van een pony, volledig “paardachtig” lichaam, rechte ruggegraat, lange nek, lange poten, gefuseerde beenderen van de poten zonder rotatie, lange neus, flexibele muil, diepe kaak. De hersenen waren een beetje groter dan bij de vroege Dinohippus. Had nog steeds enkele primitieve kenmerken zoals eenvoudige tanden en lichte gezichtsgroeven welke de latere Equussoorten verloren. Deze “eenvoudige equussoorten” diversifieerden snel in minstens 12 nieuwe soorten in 4 verschillende groepen. Gedurende de eerste grote ijstijden van het laat pleoceen (2,6 Ma) staken bepaalde equussoorten over naar de Oude Wereld. Wereldwijd nam Equus de niche over van “grote harde grassen grazende vlakteloper”.
Equus (Hippotigris) (pleistoceen). Subgenus van de moderne eentenige veervoetige grazende zebra’s.
Equus (Equus) (pleistoceen). Subgenus van de moderne eentenige veervoetige grazende paarden en ezels. (Merk op: heel af en toe wordt een paard geboren met kleine zijtenen, waaruit blijkt dat sommige paarden de genen van zijtenen behouden).
Vergelijk Equus met Hyracotherium en zie hoeveel het is veranderd. Als je bedenkt dat dieren verdeeld zijn in “soorten”, denk je dan dat Equus en Hyracotherium kunnen beschouwd worden als dezelfde “soort”? Tapirs en neushoorns:
Homagalax (vroeg-Eoceen). Erg gelijkend op het zustergenus Hyracotherium, maar de toppen waren verbonden tot ribbels dwars op de tanden. Merk op dat deze vroege perrisodactylen slechts verschilden in geringe details van de tanden.
Heptodon (laat-vroeg Eoceen). Een kleine tapirachtige met nog een verandering in de tandcusp. Splitste in twee lijnen:
Helaletes (midden eoceen) die een korte proboscis had, dan Prototapir (laat oligoceen), gelijkend op de moderne tapirs maar zonder hun flexibele snuit, dan Miotapirus (vroeg mioceen), een bijna moderne tapir met een flexibele snuit, dan Tapirus (plioceen) de moderne tapir.
Hyrachyus (laat eoceen), een tapiroïde met verhoogde schaarfunctie in zijn tanden. Leidde naar de laat-Eoceen Hyracodontiden zoals Hyracodon (neushoorn-tapiroiden, of “lopende neushoorns” die een verhoogde ontwikkeling vertonen van tanden met hoge kronen en een groter lichaam. Deze leidden naar Caenopus (vroeg-Oligoceen), een grote, niet gehoornde gegeneraliseerde neushoorn die leidde naar de moderne gehoornde neushoorns van het mioceen en plioceen. Onze levende genera verschijnen voor het eerst in het plioceen, rond 4 Ma.
Soort-soort-transities:
Paarden: Gingerich (1980) beschreef speciatie van Hyracotherium grangeri tot H. aemulor. Protero en Schoch (1989) vernoemen enkele intermediaire fossielen die de late Orohippus linken aan Mesohippus celer. MacFadden (1985) heeft vele vloeiende overgangen beschreven tussen de drietenige paarden, vooral bij de Merychippus en de verschillende hipparions. Hulbert (in Protero en Schoch, 1989) toonde aan dat Dinohippus geleidelijk overgaat in Equus langs opeenvolgende pliocene strata. Simpson (1961) beschrijft het geleidelijk verlies van de zijtenen in Pliohippus doorheen 3 opeenvolgende strata in het vroeg plioceen.
Neushoorns : Wood (1954) zei over de neushoornfossielen: “ telkens we een positief paleontologisch bewijs hebben, dan merken we een extreem gradualisme” (geciteerd in Gingerich, 1977), en Kurten (1968) beschrijft een vloeiende overgang tussen Dicerorhinus-soorten.
Minchenella of een gelijkaardige condylarth (laat paleoceen). Enkel bekend van onderkaken. Heeft een kenmerkende verbrede rand op de derde kies. De meest plausibele voorouder van de emrithopoden en anthracobuniden.
Phenacolophus (laat-Paleoceen of vroeg-Eoceen). Een vroege embrithopode (zeer vroege, licht olifantachtige condolyarthen), waarvan gedacht wordt dat ze de stamgroep zijn van alle olifanten.
Pilgrimella (vroeg eoceen). Een anthracobunide (vroege proto-olifant condylartrth), met massieve toppen op de kiezen, gelijnd in 2 transverse kammen. ° Onbenoemde soort van proto-olifant (vroeg eoceen). Recent ontdekt in Algerije. Had licht vergrote bovensnijtanden (het begin van slagtanden), en verschillende tandreducties. Had nog steeds de “normale” kiezen in plaats van de eigenaardige meervoudig gelaagde kiezen van moderne olifanten. Had het hoge voorhoofd en gepneumatiseerde (i.e. gevuld met lucht) schedelbeenderen van latere olifanten en was duidelijk een traag dier met zware beenderen. Slechts een meter hoog.
Moeritherium, Numidotherium, Barytherium (vroeg midden eoceen). Een groep gelijksoortige zeer vroege olifanten. Het is onduidelijk welke van de drie het eerst kwam. De grootte van een varken, met stevige poten, brede voeten en platte hoeven. Olifantachtig gezicht met het oog ver vooruit, en een zeer diepe kaak. Tweede snijtanden vergroot in kort slagtanden, in boven en onderkaak; kleine eerste snijtanden nog aanwezig; verlies van sommige tanden. Geen slurf.
Paleomastodon, Phiomia (vroeg oligoceen). De eerste “mastodonten”, een middelgroot dier met een slurf, lange onderkaak, en korte boven- en onderste slagtanden. Verloor eerst snijtanden en hoektanden. Kiezen hebben nog zware afgeronde toppen, met emailbanden die onregelmatig worden. Phiomia was tot 2,4 m hoog.
LACUNE: Hier is opnieuw de oligocene opening. Geen olifantenfossielen gedurende verschillende miljoenen jaren.
Gomphotherium (vroeg mioceen). Een grote uitgave van Phiomia, met aan de tanden emailranden die zeer onregelmatig worden. Twee lange rijen toppen op de tanden die dwarsribbels werden wanneer ze afsleten. Hieruit kwamen verschillende families van olifantverwanten voort die zich over heel de wereld verspreidden. Van hieraf zijn de evolutielijnen van de olifant gekend tot op het vlak van de soorten.
Olifanten
De evolutielijn van de mastodont splitste zich hier af, werd meer aangepast aan een bos loofeter niche, en gaat van Miomastodon (mioceen) en Pliomastodon (plioceen) tot Mastodon (of Mammoet”, pleistoceen).
Ondertussen werd de evolutielijn van de olifant steeds groter, zich aanpassend aan een savanne/ steppe grazende niche:
Stegotetrabelodon (laat mioceen). Een van de eerste “echte” olifanten, maar had nog steeds twee lange rijen dwarsribbels, functionele kiezen en lagere slagtanden. Andere vroege mioceen genera tonen indrukken van de toppen in platen (een modern kenmerk), met exact evenveel platen als er cusps waren. Kiezen beginnen te groeien van voor naar achter, naar voren in de kaak opschuivend gedurende de levensduur.
Primelephas (laatste mioceen). Korte onderkaak zorgt dat deze op een olifant lijkt. Vermindering en verlies van voorkiezen. Zeer veel platen op de kiezen nu; we zijn aan het bizarre systeem gekomen van de moderne olifant met een enorme multigelaagde kies die gedurende een periode functioneel is, opschuivend naar voren in de kaak.
Primelephas gomphotheroides (midden plioceen). Een latere soort die zich opsplitste in drie evolutielijnen, Loxodonta, Elephas en Mammuthus:
Loxodonta adaurora (5 Ma). Hieruit kwam de moderne olifant Loxodonta africana rond 3,5 Ma
Elephas ekorensis (5 Ma) , een vroege Aziatische olifant met nogal primitieve kiezen, duidelijk direct afgeleid uit P. gomphotheroides. Leidde rechtstreeks naar:
Elephas recki, die een zijtak had E. hydrusicus, 3,8 Ma, en dan verderging zichzelf te veranderen tot hij E. iolensis werd.
Elephas maximus, de moderne Aziatische olifant, duidelijk afgeleid uit - E. hysudricus.
Opvallend gelijkend op jonge E. hysudricus dieren. Mogelijk een geval van neotenie (waarbij “nieuwe” eigenschappen eenvoudig kenmerken zijn van jonge dieren bewaard in de volwassenheid )
Mammuthus meridionalis, duidelijk afgeleid uit P. gomphotheroiden. Verspreidde zich over het noordelijk halfrond. In Europa leidde dit tot M. armeniacus/ trogontherii, en dan tot M. primigenius. In Noord-Amerika leidde dit tot M. imperator en dan M. columbi.
Het pleistocene fossiele bestand voor olifanten is zeer goed. In het algemeen, nadat de eerste vormen van de drie moderne genera verschenen waren, vertoonden zij een zeer geleidelijke continue evolutie met bijna de helft van de aftakpunten op de evolutielijn bewaard in fossielen. Carroll bijvoorbeeld, (1988) zegt: “ Binnen het genus Elephas tonen de soorten een continue verandering gedurende een periode van 4,5 miljoen jaar… de olifanten leveren uitstekend bewijs van belangrijke morfologische verandering binnen de soorten, doorheen soorten binnen genera, en doorheen genera binnen een familie….”.
Soort-soort transities onder de olifanten:
Maglio (1973) bestudeerde pleistocene olifanten nauwkeurig. Algemeen toonde Maglio aan dat minstens 7 van de 17 olifantensoorten uit het kwartair ontstonden door geleidelijke anagenetische transities van hun voorouders. Hij zei bijvoorbeeld dat Elephas recki “ kan worden gevolgd doorheen een progressieve serie van fasen… Deze fasen lopen bijna onmerkbaar in elkaar over…. In het laat pleistoceen verschijnt een meer gevorderde olifant die ik behoud als een distincte soort, E. iolensis, enkel als een zaak van conventie. Alhoewel als een groep, het materiaal dat verwees naar E. iolensis is te onderscheiden van dat van E. recki, zijn er enkele intermediaire specimens bekend, en E. iolensis lijkt een zeer gevorderde, terminale fase in de specifieke evolutielijn van E. recki te vertegenwoordigen. Maglio documenteerde ook zeer geleidelijke overgangen tussen drie Euraziatische mammoetsoorten: Mammuthus meridionalis > M. armeniacus (of M. trogontherii) > M. primigenius. Lister (1993) analyseerde opnieuw mammoettanden en bevestigde Maglio’s schema van graduele evolutie in Europese mammoeten, en vond ook bewijs voor graduele transities in de Noord-Amerikaanse mammoeten.
Sirenes (doejongs en manatees)
Lacune: De voorouders van sirenes zijn onbekend. Geen sireneachtige fossielen zijn bekend van vóór het eoceen.
Vroeg eoceen. Fragmentaire sirenenfossielen gekend uit Hongarije
Prorastomus (midden eoceen). Een zeer primitieve sirene met een bijzonder primitieve tandenformule (waaronder de oude vijfde voorkies die alle andere zoogdieren verloren in het krijt! Betekent dit dat de sirenen zich al vroeg afsplitsten van de andere zoogdieren? De schedel is een beetje condylarthachtig. Had distincte sirene ribben. Niet voldoende van de rest van het skelet werd gevonden om te weten hoe aquatisch het dier was.
Protosiren (laat eoceen). Een sirene met over het algemeen een modern skelet, alhoewel het nog steeds de zeer primitieve tandformule had. Splitste misschien in twee overlevende evolutielijnen: 1. Doejong: Eotheroiden (laat eoceen) met een lichtjes gebogen snuit en kleine slagtanden, nog steeds met de primitieve tandformule. Hieruit kwam mogelijk Halitherium voort (oligoceen), een doejongachtige sirene met een meer gebogen snuit en langere slagtanden, en dan de bestaande doejongs, zeer gebogen snuit en grote slagtanden.
2. Manatees: sirenotherium (vroeg mioceen); Potamosiren (laat mioceen) , een manatee-achtige sirene met verlies van enkele kaaktanden; dan Ribodon (vroeg Plioceen), een manatee met continue vervanging van tanden, en dan de huidige manatees.
|
|


 7 januari 2022, 14:31
7 januari 2022, 14:31